Queen Pheromones
Scientific Beekeeping
info@scientificbeekeeping.co.uk
Chemical Communication in the Honey Bee Society.
Lack of causes
Lack of causes
HYPOPHARYNGEAL GLAND (HPG)
The hypopharangeal glands (HPG)are also found in the head under the 'forehead' or frons above the brain. The individual oval bodies are white and are set either side of a central tube. If dissecting the head of the bee this glands stretched out are longer than the bee measuring about 3cm. The secretion is clear and contains protein, fats and minerals. The outlets are into ducts from the right and left HPGs onto either side of the hypopharyngeal plate where the mouth meets the pharynx.
In queen honey bees (and drones) the HPG is vestigial (Snodgrass 1956)
The hypopharangeal glands (HPG)are also found in the head under the 'forehead' or frons above the brain. The individual oval bodies are white and are set either side of a central tube. If dissecting the head of the bee this glands stretched out are longer than the bee measuring about 3cm. The secretion is clear and contains protein, fats and minerals. The outlets are into ducts from the right and left HPGs onto either side of the hypopharyngeal plate where the mouth meets the pharynx.
In queen honey bees (and drones) the HPG is vestigial (Snodgrass 1956)
TARSAL GLANDS
The tarsal glands are also found on the ventral aspect of the feet. They produce the Footprint Hormone. They were identified by Arnhart in 1923 and are sometimes called after him. The glands consist of a single layer of glandular epithelium on the 6th tarsomere of each of the legs.
These secretions are said to inhibit queen cup construction. When a hive is crowded the queen doesnt manage to walk on the periphery of the frames so this is where swarm cells are built (see paper left).
The tarsal glands are also found on the ventral aspect of the feet. They produce the Footprint Hormone. They were identified by Arnhart in 1923 and are sometimes called after him. The glands consist of a single layer of glandular epithelium on the 6th tarsomere of each of the legs.
These secretions are said to inhibit queen cup construction. When a hive is crowded the queen doesnt manage to walk on the periphery of the frames so this is where swarm cells are built (see paper left).
DUFOUR GLANDS
The dufour glands were identifed in 1841 and used to be called the "alkaline gland".
The positional anatomy of the Dufour glands was finally established in 2004 by Martin et al (see link to left). It is a small gland associated with the venom gland, sting sheath and Koshevnikov glands (see below). it opens onto the dorsal vaginal wall close to the setosa membrane, which surrounds the sting bulb and, being hairy is an excellent vehicle for pheromone release.
To quote from the Martin et al (link to the left) re the position of the dufour gland
"Both the exits of the venom sac and Dufour gland approach the base of the sting bulb. However, where the venom sac opens directly into the base of the sting bulb the Dufour gland narrows before exiting between the sting lancets. The exit is very narrow and indistinct, which explains the confusion in the literature and inability to observe the exit using a binocular microscope (×60). In A. mellifera ... the Dufour gland exit is in the same position in both workers and queens"
The dufour glands were identifed in 1841 and used to be called the "alkaline gland".
The positional anatomy of the Dufour glands was finally established in 2004 by Martin et al (see link to left). It is a small gland associated with the venom gland, sting sheath and Koshevnikov glands (see below). it opens onto the dorsal vaginal wall close to the setosa membrane, which surrounds the sting bulb and, being hairy is an excellent vehicle for pheromone release.
To quote from the Martin et al (link to the left) re the position of the dufour gland
"Both the exits of the venom sac and Dufour gland approach the base of the sting bulb. However, where the venom sac opens directly into the base of the sting bulb the Dufour gland narrows before exiting between the sting lancets. The exit is very narrow and indistinct, which explains the confusion in the literature and inability to observe the exit using a binocular microscope (×60). In A. mellifera ... the Dufour gland exit is in the same position in both workers and queens"
It is apparent that the Dufour gland chemicals are a fertility signal. In workers these are long chain hydrocarbons but in queens there is also found waxlike esters and these esters are closley correlated to ovarian development.
Esters found in Dufour Gland Pheromone: (Katzav 2001)
Tetradecyl dodecanoate
Tetradecyl (Z)-9-tetradecenoate
Tetradecyl tetradecanoate
Tetradecyl (Z)-9-hexadecenoate + Tetradecyl (Z)-11-hexadecenoatea (trace)
Tetradecyl hexadecanoate + Hexadecyl tetradecanoate
Tetradecyl (Z)-9-octadecenoate
(Z)-9-hexadecenyl hexadecanoate
Hexadecyl hexadecanoate
Octadecyl tetradecanoate
(Z)-9-hexadecenyl (Z)-9-octadecenoate
Octadecyl hexadecanoate
Queens with multiple matings were found to be more attractive than those with a single drone mate.
An earlier hypothesis that the Dufour gland chemicals serve as an egg marking pheromone was disproved.
Esters found in Dufour Gland Pheromone: (Katzav 2001)
Tetradecyl dodecanoate
Tetradecyl (Z)-9-tetradecenoate
Tetradecyl tetradecanoate
Tetradecyl (Z)-9-hexadecenoate + Tetradecyl (Z)-11-hexadecenoatea (trace)
Tetradecyl hexadecanoate + Hexadecyl tetradecanoate
Tetradecyl (Z)-9-octadecenoate
(Z)-9-hexadecenyl hexadecanoate
Hexadecyl hexadecanoate
Octadecyl tetradecanoate
(Z)-9-hexadecenyl (Z)-9-octadecenoate
Octadecyl hexadecanoate
Queens with multiple matings were found to be more attractive than those with a single drone mate.
An earlier hypothesis that the Dufour gland chemicals serve as an egg marking pheromone was disproved.
KOSCHEVNOKOV GLAND (spelt various ways in different texts)
The Koschevnokov Gland (KG) can be seen in the diagram above. It is made of many smaller glandular units, each with a secretory cell and duct cell connected to the epidermis. The secretion is released onto the setosa membrane.
For completeness the following are found in the queen's KG pheromones:
1,1,3-Trimethyl cyclopentane
5,5-Dðyl-2-hexene
3,3-Dimethyl-hexane
Octenal
Methyl cyclcdecane
p-menthane-9-01
4,5-Dimethyl-nonane
2-Propyl-1-heptanol
4,6,8-Trimethyl-I-nonene
Nonanoic acid
Decanoic acid
1,12-Tridecadiene
1.1 I -Dodecadiene
Cyclohexyl hexenol
Ethyl decanoate
2-Methyl-I-dodecanol
Hexadecane
Ethyl dodecanoate
Dodecyl acetate
6-Cyclohexyl undecane
Hexadecanoic acid
Ethyl tetradecanoate
Methylester 2-methyl hexadecanoate
2-(Hexadccyl oxy)-ethanol
2,6,10,15-Tetramethyl-heptadecane
1-Dotriacontanol
1,7-Pentatriacontene
3,5,24-Trimethyl-tetracontane
The gland starts to degenerate after 1 year and contributes to the loss of queen signal. It has a primer function.
The Koschevnokov Gland (KG) can be seen in the diagram above. It is made of many smaller glandular units, each with a secretory cell and duct cell connected to the epidermis. The secretion is released onto the setosa membrane.
For completeness the following are found in the queen's KG pheromones:
1,1,3-Trimethyl cyclopentane
5,5-Dðyl-2-hexene
3,3-Dimethyl-hexane
Octenal
Methyl cyclcdecane
p-menthane-9-01
4,5-Dimethyl-nonane
2-Propyl-1-heptanol
4,6,8-Trimethyl-I-nonene
Nonanoic acid
Decanoic acid
1,12-Tridecadiene
1.1 I -Dodecadiene
Cyclohexyl hexenol
Ethyl decanoate
2-Methyl-I-dodecanol
Hexadecane
Ethyl dodecanoate
Dodecyl acetate
6-Cyclohexyl undecane
Hexadecanoic acid
Ethyl tetradecanoate
Methylester 2-methyl hexadecanoate
2-(Hexadccyl oxy)-ethanol
2,6,10,15-Tetramethyl-heptadecane
1-Dotriacontanol
1,7-Pentatriacontene
3,5,24-Trimethyl-tetracontane
The gland starts to degenerate after 1 year and contributes to the loss of queen signal. It has a primer function.
TERGAL GLANDS
Tergal glands have a releaser effect on the workers of a honey bee colony. Worker bees can often be found encircling the queen. They can be seen to antennate the queen and lick her abdomen. The pheromones are of low volatility and can only be picked up when very close to the queen.
Decyl decanoate and longer chain-length esters of decanoic acid are dominant components of the pheromone from the abdominal tergite glands of virgin honeybee queens, 3–10 days old. These esters are present in very small amounts in queens which were 0–2 days old. The cuticular lipids of honeybee queens were predominantly n-alkanes and alkenes with and being major components.
hexadecanoic acid methyl ester (methyl palmitate)
(Z)-9-hexadecenoic acid (palmitoleic acid)
hexadecanoic acid (palmitic acid)
(Z)-9- octadecenoic acid (oleic acid) [dominant in virgin queens]
(E)-11-octadecenoic acid (vaccenic acid)
octadecanoic acid methyl ester (methyl stearate )
octadecanoic acid (stearic acid)
decanoic acid (lauric a c i d ) ,
decyl decanoate,
n-alkanes (major part n-heptacosane)
alkenes (major part hentriacontene) [Only produced after mating]
(Al-Qarni et al. 2005)
Queens are also thought to produce a close-range sex attractant from the tergal glands. The active compound has been proposed to be decyl decanoate (Espelie et al., 1990).
Smith et al. (1993) demonstrated in their experiments that the production of tergal gland alkenes is stimulated by natural mating and not by experimental insemination.
They said that It has long been recognized in the beekeeping industry that instrumentally inseminated queens are not as pro-ductive as naturally mated queens. Problems are observed with initial introduction and acceptance of the inseminated queens, rapid replacement of the introduced inseminated queen by a queen raised from her eggs and decreased brood production by inseminated queens. The tergal gland alkenes may play a key role in the care and acceptance of the queen and her eggs by worker bees in the hive (Smith et al., 1993)
The above was refuted in later research by Al-Qarni et al 2003.
Tergal glands have a releaser effect on the workers of a honey bee colony. Worker bees can often be found encircling the queen. They can be seen to antennate the queen and lick her abdomen. The pheromones are of low volatility and can only be picked up when very close to the queen.
Decyl decanoate and longer chain-length esters of decanoic acid are dominant components of the pheromone from the abdominal tergite glands of virgin honeybee queens, 3–10 days old. These esters are present in very small amounts in queens which were 0–2 days old. The cuticular lipids of honeybee queens were predominantly n-alkanes and alkenes with and being major components.
hexadecanoic acid methyl ester (methyl palmitate)
(Z)-9-hexadecenoic acid (palmitoleic acid)
hexadecanoic acid (palmitic acid)
(Z)-9- octadecenoic acid (oleic acid) [dominant in virgin queens]
(E)-11-octadecenoic acid (vaccenic acid)
octadecanoic acid methyl ester (methyl stearate )
octadecanoic acid (stearic acid)
decanoic acid (lauric a c i d ) ,
decyl decanoate,
n-alkanes (major part n-heptacosane)
alkenes (major part hentriacontene) [Only produced after mating]
(Al-Qarni et al. 2005)
Queens are also thought to produce a close-range sex attractant from the tergal glands. The active compound has been proposed to be decyl decanoate (Espelie et al., 1990).
Smith et al. (1993) demonstrated in their experiments that the production of tergal gland alkenes is stimulated by natural mating and not by experimental insemination.
They said that It has long been recognized in the beekeeping industry that instrumentally inseminated queens are not as pro-ductive as naturally mated queens. Problems are observed with initial introduction and acceptance of the inseminated queens, rapid replacement of the introduced inseminated queen by a queen raised from her eggs and decreased brood production by inseminated queens. The tergal gland alkenes may play a key role in the care and acceptance of the queen and her eggs by worker bees in the hive (Smith et al., 1993)
The above was refuted in later research by Al-Qarni et al 2003.
Adapted from Goodman, L. (2003) Form and Function in the Honey Bee. Cardiff, UK: IBRA
MANDIBULAR GLAND (MG)
Dr Colin Butler did much research in the 1950's, 60s and 70's on bee behaviour which identified, amongst other things, that queen bees produce Queen Mandibular Pheomone and the actions caused either through its presence or absence. It behaves as a primer within her colony, inhibiting worker ovary development and keeping a swarm together during swarming. It also acts as a primer in an attractant for drones when flying in a drone congregation area.
The MGs are located in the 'cheeks' or Gena of the bee's face. Their secretions appear on the inner surface of the mandibles. The main chemical the queen produces is 9-oxodec-2-enoic acid (9-ODA).
There appear to be three different patterns in queen MP according to reproductive state but research so far is inconclusive. It seems to be possible to postulate that:
(1) Newly emerged virgin premating queens presented a weak signal, with oleic acid (OLA) as the main component,
(2) More mature virgin queens ready for mating have an intensified signal, mainly consisting of 9-oxodec-2-enoic acid (9-ODA) along with OLA and small amounts of 9-hydroxydec-2-enoic acid (9-HDA), and
(3) postmating dominant queens exhibited a strong signal with high concentrations of 9-ODA combined with medium proportions of 9-HDA, less OLA, and small amounts of oxygenated aromates.
(4)Some research has suggested that oxygenated aromates, especially methyi p-hydroxybenzoate (HOB) and the late appearing 4-hydroxy-3-methoxy-phenylethanol (HVA), could be the typical signal of old egg-laying and dominant queens. Perhaps this is the signal the worker bees receive to tell them they need to replace their queen.
What appears to be clear is that 9-HDA is oxidised to become 9-ODA. Young virgin queens appear unable to oxidise 9-ODA. It is 9-ODA in particular that prevents new queen cells and laying workers.
It is therefore these five components;
OLA= oleic acid [just emerged queen]
9-HDA= 9-hydroxydec-2-enoic acid [more mature virgin queens]
9-ODA = 9-oxodec-2-enoic acid [Mated Queen in full lay]
HOB= methyi p-hydroxybenzoate [Older laying Queen]
HVA= 4-hydroxy-3-methoxy-phenylethanol [Older, possibly failing queen])
that make up the principle QMP throuout her life.
Dr Colin Butler did much research in the 1950's, 60s and 70's on bee behaviour which identified, amongst other things, that queen bees produce Queen Mandibular Pheomone and the actions caused either through its presence or absence. It behaves as a primer within her colony, inhibiting worker ovary development and keeping a swarm together during swarming. It also acts as a primer in an attractant for drones when flying in a drone congregation area.
The MGs are located in the 'cheeks' or Gena of the bee's face. Their secretions appear on the inner surface of the mandibles. The main chemical the queen produces is 9-oxodec-2-enoic acid (9-ODA).
There appear to be three different patterns in queen MP according to reproductive state but research so far is inconclusive. It seems to be possible to postulate that:
(1) Newly emerged virgin premating queens presented a weak signal, with oleic acid (OLA) as the main component,
(2) More mature virgin queens ready for mating have an intensified signal, mainly consisting of 9-oxodec-2-enoic acid (9-ODA) along with OLA and small amounts of 9-hydroxydec-2-enoic acid (9-HDA), and
(3) postmating dominant queens exhibited a strong signal with high concentrations of 9-ODA combined with medium proportions of 9-HDA, less OLA, and small amounts of oxygenated aromates.
(4)Some research has suggested that oxygenated aromates, especially methyi p-hydroxybenzoate (HOB) and the late appearing 4-hydroxy-3-methoxy-phenylethanol (HVA), could be the typical signal of old egg-laying and dominant queens. Perhaps this is the signal the worker bees receive to tell them they need to replace their queen.
What appears to be clear is that 9-HDA is oxidised to become 9-ODA. Young virgin queens appear unable to oxidise 9-ODA. It is 9-ODA in particular that prevents new queen cells and laying workers.
It is therefore these five components;
OLA= oleic acid [just emerged queen]
9-HDA= 9-hydroxydec-2-enoic acid [more mature virgin queens]
9-ODA = 9-oxodec-2-enoic acid [Mated Queen in full lay]
HOB= methyi p-hydroxybenzoate [Older laying Queen]
HVA= 4-hydroxy-3-methoxy-phenylethanol [Older, possibly failing queen])
that make up the principle QMP throuout her life.
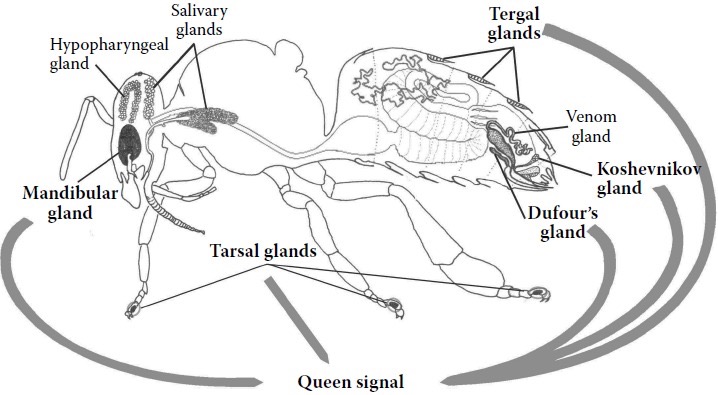
In the often used image above we can identify all the sources of pheromones and other enzymes that arise from the queen bee. They will alter according to her fertility status. There is a separate page for Queen Retinue Pheromones, many of which will be found referenced this page. Here I have listed all pheromones I have found scientifically established to be present in the Apis mellifera queen. Not all chemicals have an established purpose to date.
The diagram above gives a simple impression of the affects that the queen pheromones, also known as Queen Signal, have on the colony.
The primary functions are considered to have primer effects. The colony, like all organisms, needs to remain in a state of homeostasis. A colony is considered to be a superorganism. Key to the colony is a fecund queen. If she is killed, injured to prevent egg laying or becomes sterile the QS diminishes and queen rearing or laying works will ensue.
The diagram above gives a simple impression of the affects that the queen pheromones, also known as Queen Signal, have on the colony.
The primary functions are considered to have primer effects. The colony, like all organisms, needs to remain in a state of homeostasis. A colony is considered to be a superorganism. Key to the colony is a fecund queen. If she is killed, injured to prevent egg laying or becomes sterile the QS diminishes and queen rearing or laying works will ensue.
STING GLANDS (QUEEN)
Young queen honeybees use their venoms only for stinging other queens and possess a venom only half as lethal to mice as worker venom, and by the time queens are 1–2 years of age their venom has become essentially inactive.
The sting glands of bees produce two principle chemicals, melittin and phospholipase A2. In an article by Danneels et al (see link left) many more substances were identified occurring by spectroscopy varing in presence in queens, summer workers and winter workers. The queens were found to lack 6 of the 34 venom toxins present in worker bees at some time in the year. Queens were also found to have 2 unique toxins: serine proteinase stubble and antithrombin-III
Young queen honeybees use their venoms only for stinging other queens and possess a venom only half as lethal to mice as worker venom, and by the time queens are 1–2 years of age their venom has become essentially inactive.
The sting glands of bees produce two principle chemicals, melittin and phospholipase A2. In an article by Danneels et al (see link left) many more substances were identified occurring by spectroscopy varing in presence in queens, summer workers and winter workers. The queens were found to lack 6 of the 34 venom toxins present in worker bees at some time in the year. Queens were also found to have 2 unique toxins: serine proteinase stubble and antithrombin-III
NB
There are other enzymes produced by the queen bee, which appear to keep her reproductive organs in optimal work order: Catalase (CAT), glutathione $S$-transferase (GST) and superoxide dismutase (SOD). It is believed that they play a role in the long-term protection of the spermatozoa from oxidative stress.
There are other enzymes produced by the queen bee, which appear to keep her reproductive organs in optimal work order: Catalase (CAT), glutathione $S$-transferase (GST) and superoxide dismutase (SOD). It is believed that they play a role in the long-term protection of the spermatozoa from oxidative stress.